غشای سلول را بیشتر بشناسیم!

مقدمه
در مقاله سلول، ما به طور مختصر درباره سلول و اجرای آن صحبت کردیم. مانند هر قلمرو دیگری، سلول نیز دارای دیواره ای است که مرزهای آن را از محیطش مجرا می کند. اما به عنوان یک بافت زنده و فعال، دیواره های محافظ سلول نمی تواند غیر فعال و ثابت باشد و قطعاً نیاز دارد تا وظایف مختلف در جهت حفظ و تداوم حیات سلول و مهم تر از آن، ارتباط سلول با اطراف را به دوش بکشد. بیایید ببینیم این غشای فعال دقیقا چه ساختاری و عملکردی دارد و این ساختارها چگونه با عملکردهای مختلف آن تناسب یافته است.
بخش اول: غشای سلول و تاریخچه کشف آن
چه می آموزیم؟
- قبل از هر چیز، بیایید ببینیم منظور از غشای سلول چیست و چه کسانی آن را کشف کرده اند.
غشای سلولی چیست؟
منظور از غشای سلول، دیواره پوشاننده هر سلول است که دور تا دور سلول را، بدون کوچکترین شکافی، احاطه کرده و چند وظیفه اصلی و مشترک بین انواع سلول ها دارد:
- سلول های را کنار هم نگه می دارد.
- ورود و خروج مواد بین دو سمت غشا را کنترل می کند (انتشار، اسمزی، انتقال فعال)
- از سلول در برابر عوامل خارجی محافطت می کند.
- به سلول اجازه می دهد تا اطراف خود را بشناسد و شناخته شود.
- به سلول های دیگر یا به مولکول ها می چسبد.
- محلی برای واکنش های بیوشیمیایی است (آنزیمی، مناطق واکنشی)
در مقاله سلول، خلاصه ای از ساختار سلول و بخش های مختلف آن، به ویژه غشا را بررسی کردیم و نشان دادیم که غشا یک ساختار دو لایه از مولکول های فسفولیپید است که در موازات هم قرار گرفته و یک مرکز آب گریز و دو لایه بیرونی آب دوست را تشکیل داده اند.
در بخش های بعدی، هر یک از ساختارهای موجود در غشا را به تفصیل بررسی خواهیم کرد.
تاریخچه کشف غشای سلول
پیش از کشف سلول و غشای آن، کشف دستگاه هایی که به کمک آن ها بتوان این ساختار ها را مشاهده کرد، اهمیت می یابد. اولین با در تاریخ در 1590، زاخاریس جانسن (Zacharias Jansen)، که یک تولید کننده عینک آلمانی بود، یک میکروسکوپ نوری پروتوتایپ را اختراع کرد (1). نوزده سال پس از او، گالیله گالیله (Galileo Galilei) که یک دانشمند ایتالیایی بود، یک میکروسکوپ نوری ساخت (2). علی رغم اختراع میکروسکوپ، مشاهده سلول در زیر میکروسکوپ سال ها بعد برای اولین بار انجام شد. برای شناخت غشای سلول، تحقیقات وسیعی در قرون 16 تا 19 انجام گرفت. دانشمند بزرگ که پدر علم اپتیک نام گرفت، رابرت هوک (Robert Hooke) نام داشت. او با میکروسکوپ گیاهان، مخمرها، دانه های ماسه و کک را مشاهده کرد. او کسی بود که سلول را کشف و آن را نامگذاری کرد. او سهم قابل توجهی در رشد دانش در زمینه های فیزیک، شیمی، هواشناسی، جغرافیا و زیست شناسی داشت (3). در اوایل دهه 1660، کریستوفر رن (Christopher Wren)، توماس ویلیس (Thomas Willis) و چند محقق دیگر به همراه رابرت هوک در گروه آزمایشی آکسفورداولین مشاهدات خود را به وسیله میکروسکوپ نوری انجام دادند. در سال 1665، پس از آن که هوک به لندن مهاجرت کرد، زمانی که داشت بر روی یک تکه قارچ زیر میکروسکوپ نوری آزمایش می کرد، شکاف هایی مشاهده کرد و آن ها را «سلول» (Cellula) نامید، که به معنای فضا (Space) بود (4). چهار سال بعد، دو مقاله مهم توسط گیاه شناس انگلیسی نهمیا گرو (Nehemiah Grew) و آناتومیست ایتالیایی مارچلو مالپیگی (Marcello Malpighi) منتشر شد. این دو دانشمند، بدون اینکه همدیگر را بشناسند، ریز آناتومی گیاهان را شرح داده بودند. مالپیگی همچنین ریز آناتومی سلول های جانوری را نیز شرح داده بود. از آنجایی که ساختارهای سلولی گیاهان شکل اولیه تری نسبت به سلول های جانوری داشت، اولین مطالعات بر روی گیاهان انجام گرفت (5).

رابرت هوک (1635-1703 میلادی)
در سال 1672 گرو، «توده جباب ها» در ارتباط با پارانشیم گیاهی را شرح داد. در سال 1682 او گزارش کرد که غشای سلول شبیه یک ساختار توری مانند است. در مشاهدات گرو و در همان دوران، گزارش شد که دیواره سلول های گیاهی یک لایه کاملا یکپارچه است که سلول را در بر گرفته است.
در دومین ادیت از کتاب «فلسفه جانورشناسی» که در دهه 1800 توسط لامارک (Lamarck) نوشته شد، یک بخش به بافت سلولی اختصتص داده شد. در این بخش لامارک نوشت: «غشاها که سالهاست به عنوان پوششی برای مغز، اعصاب، همه انواع رگ ها، غدد، ارگان ها، عضلات و تارهای عضلانی و حتی پوست شناخته شده اند، در واقع یکی از تولیدات بافت سلولی هستند» (6).
در سال های بعد، نویسندگانی نیز یافت شدند که وجود غشا در سلول را انکار کردند. در 1857، اولین فرد از این گروه، به نام فرانتس فون لیدیگ (Franz Von Leydig) که راجب سلول و ساختارهای سلولی مطالعه می کرد، چنین شرح داد که سلول به عنوان یک ماده نرم با یک هسته در میانه آن، توسطی پوششی در اطاف احاطه شده است. اما چنین تصور می کرد که غشای سلول یک ساختار ثانویه است که در اثر سفت شدن سطح سلول بوجود آمده است (7).
در نیمه دوم قرن نوزده، دانشمندان آزمایشی برای تراوایی غشا انجام دادند. آنان مشاهده کردند که زمانی که سلول گیاهی را در مجاورت محلول هایپرتونیک (با غلظت بالا) قرار می دهند، سلول ها منقبض شده و پروتوپلاسم (سیتوپلاسمی سلول گیاهی) از دیواره به سمت بیرون سلول جابجا می شود. به طور معکوس، زمانی که سلول در مجاورت محلول هیپوتونیک (با غلظت پایین) قرار می گیرد، سلول متورم شده و تا مرز ترکیدن پیش می رود. بعدها، در سال 1877، پففر (Pfeffer) گیاه شناس، تحت تأثیر یافته های فیزیو-شیمی قرار گرفت و پیشنهاد کرد که یک غشای پلاسمایی یا پوستی لایه بیرونی پروتوپلاسم را می پوشاند (8). او همچنین کشف کرد که این سد دفاعی پوست مانند به آب به صورت انتخابی اجازه ورود به سلول را می دهد، اما عبور نمک و قند از محدود است (9).
در سال 1904 ناتانسون (Nathanson) نظریه غشای موزائیکی را مطرح کرد. بر اساس فرضیه او، غشای سلول به صورت بخش های موزاییک مانند است و دارای مسیرهایی برای عبور مواد از بین موزائیک هاست (10). در سال 1925، اورت گورتر (Evert Gorter ) و فرانسیس گرندل (Francoise Grendel) لیپیدها را از گلبول های قرمز جدا کردند. آن ها علاوه بر کشف ساختار لیپیدی غشا، نشان دادن که مولکول ها غشا می توانند یک یا دو لایه باشند. نظریه این دو در آن زمان مفهوم دولایه لیپیدی را بر سر زبان ها انداخت. همچنین این نظریه یک سرنخ مهم برای شناختن ساختار مولکولی غشا در اختیار دانشمندان گذاشت (11).
بالاخره بین سال های 1920 تا 1930 ضخامت غشای سلول اندازه گیری شد و نشان داده شد که غشا یک ساختار لیپیدی خالص نیست، بلکه ساختارهای پروتئینی فراوانی درون آن قرار گرفته است. در سال 1935 نظریه ای مبنی بر مرکز لیپیدی غیرقطبی بین دولایه آمفی پاتیک غشا مطرح شد.
علی رغم اینکه میکروسکوپ الکترونی در سال 1930 کشف شد، تا دهه 1950 که امکان تصویر برداری با وضوح بالا وجود نداشت، با این وسیله بر روی غشای سلول مطالعه نشده بود. برش عرضی غشا، یک ساختار سه لایه را نشان داد که با نام «راه آهن» شناخته شد. در این مشاهده که بعدها «واحد غشایی» نامیده شد، دولایه چگال با یک لایه سبک بین آن ها نشان داده شد. در آن زمان هنوز مشخص نبود در هر لایه چه مولکول هایی وجود دارد. به طور ابتدایی در سال 1959، پیشنهاد شد که لایه چگال از جنس پروتئین و لایه سبک از جنس چربی است که با دانش امروز ما اکنون می دانیم که این نظریه صحیح نیست.
در سال 1964، بردی و ترامس (Brady & Trams) مدلی تحت عنوان موزائیک سیال را عنوان کردند که تا امروز نیز مورد پذیرش است. در این مدل با دانش امروزی، واژه موزائیک به ساختارهای مختلف مولکولی غشا، شامل فسفولیپیدها، پروتئین ها و کربوهیدرات ها اشاره می کند و واژه سیال، نشان می دهد که بیشتر این ساختارها قابلیت جابجایی دارند (12).
در دهه 1970 این مدل توسعه یافت و اساس آن تا امروز بدن تغییر مانده است. مطالعات بعدی بر سوءعملکردهای غشا، سیگنالینگ های غشایی، انتقال در عرض غشا، تغییر شکل آن و بیوسنتز غشا متمرکز شدند.
بخش دوم: مولکول های ساختاری غشا
چه می آموزیم؟
پس از نگاهی گذرا به غشای سلول و تاریخچه کشف آن، بیایید ساختار غشا را از نزدیک تر ببینیم.
ساختار مولکولی غشا
تقریباً غشای همه سلول های یوکاریوتی یک ساختار مولکولی دولایه است. هر لایه از این غشا، از کنار هم قرار گرفتن مولکول های گلیسروفسفولیپید ساخته شده است. در زیر می توانید یک مولکول گلیسروفسفولیپید و قرار گیری آن در غشای دولایه را مشاهده کنید.
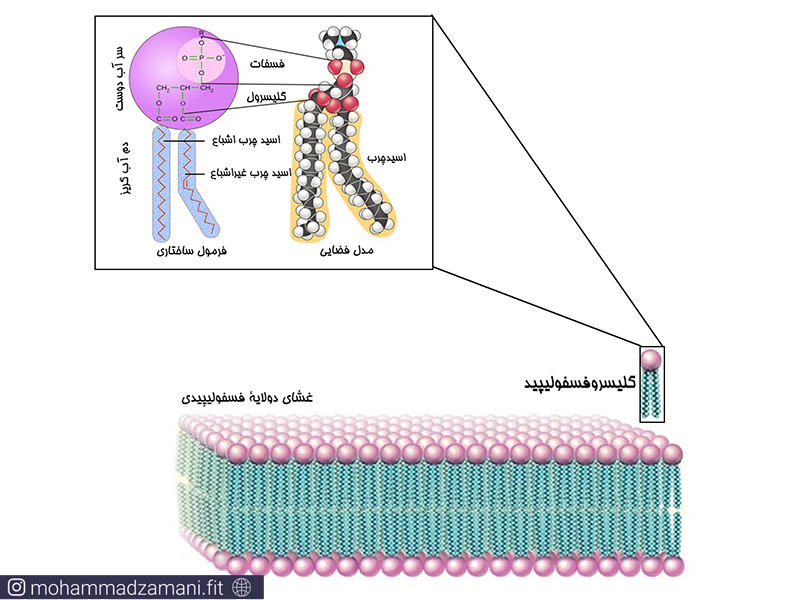
همانطور که در شکل نیز مشخص است، نحوه قرار گیری مولکول های فسفولیپید در غشا، به نحوی است که دم های لیپیدی آب گریز در مجاورت هم و سرهای فسفری آب دوست در دوسمت غشا قرار گرفته اند. این نحوه قرارگیری باعث شده است که در میانه غشا لایه ای مقاوم ایجاد شود که مانع از عبور آب و مواد محلول در آن در عرض غشا شود. این مورد اولین مکانیسم ساختاری دفاعی سلول در برابر هر آن چیزی است که در خارج از مرزهای سلول قرار دارد.
ساختارهای پروتئینی در غشا
غشای سلول ها تنها از فسفولیپید ساخته نشده است. بلکه مولکول های پروتئینی متعددی در لابلای این پوشش دولایه فسفولیپیدی پراکنده شده اند که می توانند ساختارهای متنوع و عملکردهای منحصر به فردی در سلول داشته باشند.
پروتئین های موجود در غشا یا کاملا در داخل ساختار دولایه قرار گرفته اند که در این صورت به آن ها پروتئین سرتاسری می گویند، یا در یک سمت غشا و تنها در ارتباط با مولکول ها فسفری سطح خارجی یا داخلی غشا هستند که در این صورت به آن ها پروتئین های محیطی گفته می شود..
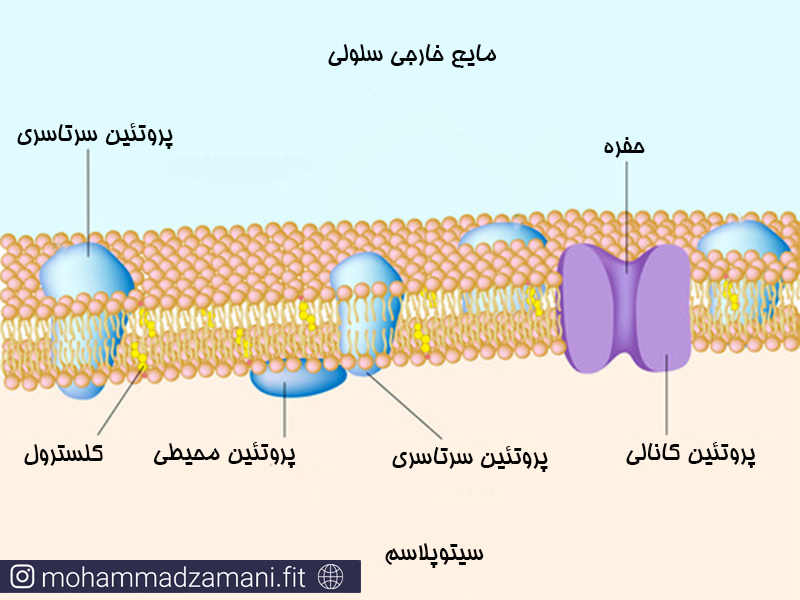
پروتئین ها در غشا وظایف متعددی به عهده دارند که اصلی ترین آن ها شامل موارد زیر است:
انتقال دهنده
همانطور که گفته شد غشای فسفولیپیدی دولایه در مقابل بسیاری از مواد نفوذناپذیر است. اما سلول برای حفظ حیاط خود نیاز با ارتباط و دادوستد با بیرون دارند. یکی از این روش های ارتباطی وجود پروتئین های سرتاسری در غشا است که نقش انتقال مواد در عرض غشا را به عهده دارند. در بخشی مجزا، انواع انتقال و انتقال دهنده در عرض غشا را بررسی خواهیم کرد.
دریافت کننده سیگنال های شیمیایی از خارج از سلول
به این نوع پروتئین ها، معمولاً «گیرنده» گفته می شود. گیرنده ها، پیام های شیمیایی، مانند هورمون ها را از خارج سلول دریافت می کنند و در ارتباط با پروتئین های دیگر، پاسخ لازم توسط سلول را تحریک می کنند.
تبدیل سیگنال های شیمیایی به عملکرد داخل سلولی
پروتئین هایی که چنین عملکردی دارند، «آنزیم» نامیده می شوند. ویژگی منحصر به فرد این پروتئین ها، ایجاد پاسخ مستقیم در اثر تحرک است.
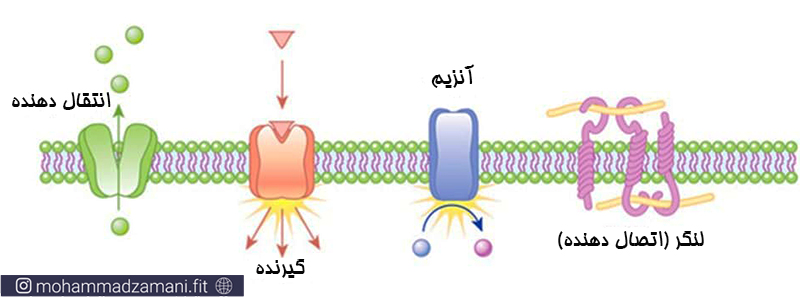
ایجاد لنگر اتصالی
این نوع پروتئین های سرتاسری وظیفه اتصال سلول به سلول مجاور یا هر نوع بافت دیگر در اطراف را بر عهده دارند و معمولاً در داخل غشا، به اسکلت سلولی و در در سمت بیرون غشا، به پروتئین همسان خود از سلول دیگر یا پروتئین های ساختاری بافتی بین سلولی، مانند کلاژن متصل هستند.
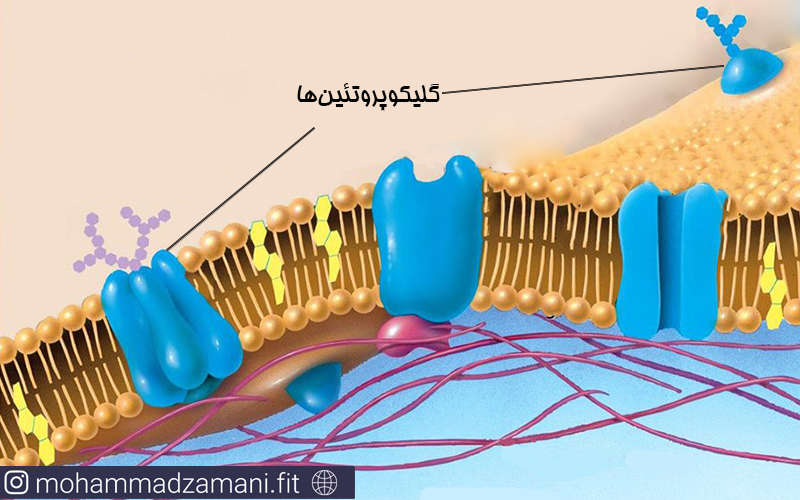
ساختارهای گلیکوپروتئینی
گلیکوپروتئین ها که در سطح غشای دولایه قرار می گیرند، در واقع ترکیبات کربوهیدراتی متصل به یکی از پروتئنی های غشا هستند. خاصیت آب دوستی این ساختارها به آن ها اجازه می دهد تا در محیط آبی به وظایف خود عمل کنند که مهم ترین آن ها شناساندن سلول به سلول های اطراف و اتصال به مولکول های دیگر برای ایفای نقش های متفاوت است. گلیکوپروتئین ها همچنین برای اتصال متقابل سلول ها و پروتئین ها (مانند کلاژن) و افزایش قدرت و پایداری سلول مهم هستند.
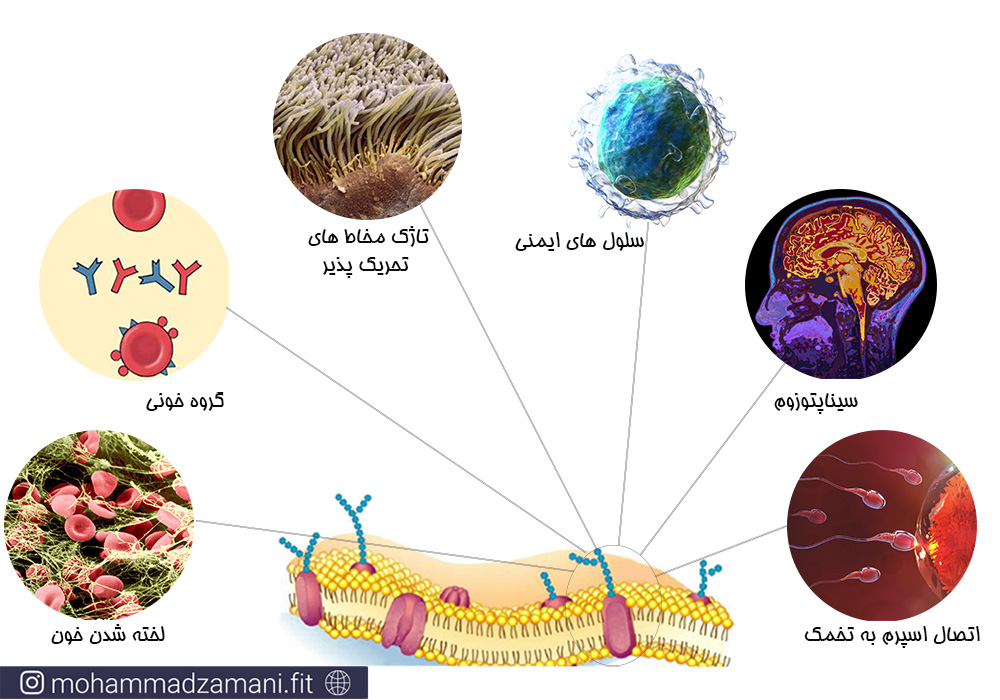
گلیکوپروتئین ها یا همان پروتئین های گلیکوزیله تنها برای ارتباطات بین سلولی مهم نیستند؛ آن ها همچنین به سیستم های ارگانی مختلف کمک می کند تا باهم ارتباط برقرار کنند. گلیکوپروتئین ها در ماده خاکستری مغز یافت می شوند و ارتباط آکسون ها و سیناپتوزوم ها را میسر می سازند.
گلیکوپروتئین ها در برخی از سلول ها عملکرد ویژه تری نیز دارند. به عنوان مثال لخته شدن خون، با کمک گلیکوپروتئین های پروترومبین، ترومبین و فیبرینوژن انجام می شود.
علاوه بر این موارد، گلیکوپروتئین های غشا در برخی سلول ها نقش شناسنامه برای سلول ایفا می کنند. گروه های مختلف خونی به روش دسته بندی MN به وسیله دو نوع متفاوت از گلیکوپروتئین غشایی گلیکوفورین A انجام می شود. همچنین دسته بندی رایج گروه های خونی ABO، بر اساس وجود گلیکوپروتئین های مختلف در غشای گلبول های قرمز خون انجام می شود.
گلیکوپروتئین های غشا همچنین برای تولید مثل نیز ضروری هستند. زیرا اسپرم از طریق همین گلیکوپروتئین ها به سلول تخمک متصل شده و لقاح انجام می شود.
تاژک هایی که در مخاط های تحریک پذیر یافت می شوند، مانند مخاط مسیرهای تنفسی و دستگاه گوارش، در واقع همان گلیکوپروتئین های غشایی هستند.
سلول های ایمنی نیز برای فعالیت محتاج به گلیکوپروتئین های غشایی خود هستند. گلیکوپروتئین های غشایی اسن سلول های ایمنی، آنتی ژن هایی که سلول باید به آن متصل شود را تشخیص می دهند. برای مثال سلول های B و سلول های T دارای این گلیکوپروتئین ها در سطح غشای خود هستند.
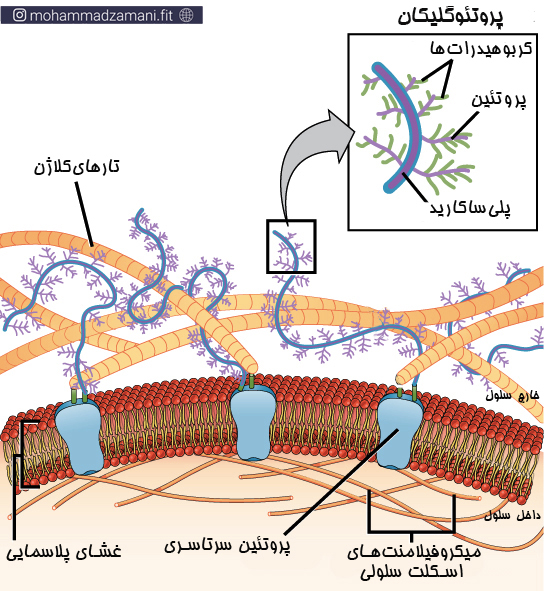
نوع دیگر ساختار گلیکوپروتئین های غشایی، مولکولی به نام «پروتئوگلیکان» است. پروتئوگلیکان ها مؤلفه های اصلی سازنده ماتریکس بین سلولی هستند؛ یک ماده پر کننده فضای بین سلولی در موجودات زنده. پروتئوگلیکان ها همچنین به کاتیون های مختلف و مولکول های آب متصل می شوند و حرکت مولکول های مختلف درون ماتریکس را تنظیم می کنند (13).
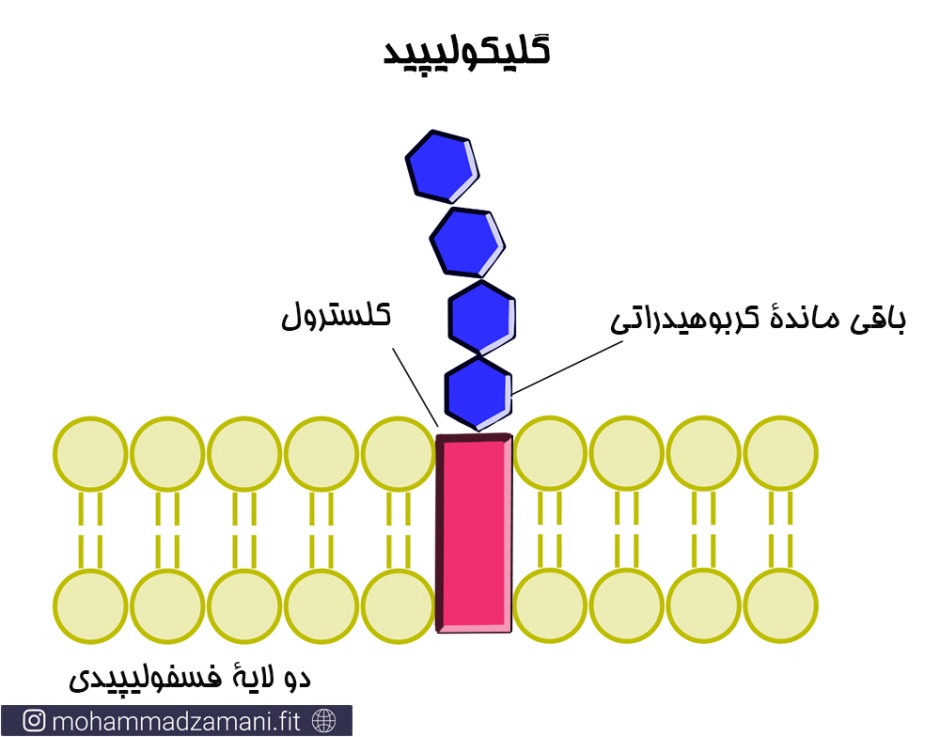
ساختارهای گلیکولیپیدی
گلیکولیپیدها ترکیبات کربوهیدراتی هستند که به مولکول های لیپیدی غشا، شامل اسفینگولیپیدها و گروه های گلیسرول، در سطح خارج سلولی متصل می شوند. از نظر عملکرد شباهت های زیادی بین گلیکوپروتئین و گلیکولیپید وجود دارد. زیرا گلیکولیپیدها نیز ساختارهای در جهت برقراری ارتباط بین سلول ها هستند. علاوه بر آن، این مولکول ها می توانند به پایداری غشا نیز کمک کنند.
گلیکولیپیدها همچنین می توانند به عنوان گیرنده ویروس ها و دیگر عوامل پاتوژن عمل کنند و تسهیل کننده ورود آن ها به سلول باشند. در سلول های جانوری گلیکولیپیدها دو دسته اصلی دارند که شامل گانگلیوزیدها و سربروزیدها هستند.
همانند گلیکوپروتئین ها، گلیکولیپیدها نیز ساختارهایی هستند که در تعیین نوع گروه خونی اثر دارند. در واقع انواع گلیکولیپید و گلیکوپروتئینی که در سطح سلول های قرمز خون وجود دارد، تعیین کننده گروه خونی هر فرد است.
سایر ساختارهای مولکولی غشا
از دیگر مولکول هایی که به صورت پراکنده ممکن است در ساختار غشا وجود داشته باشد، مولکول های کلسترول هستند که اگرچه مقدار آن ها از گلیسروفسفولیپیدها کمتر است، اما حدود 20 درصد از لیپیدهای غشای پلاسمایی سلول های حیوانی را تشکیل می دهند. کلسترول به تنظیم سفتی غشاها کمک می کند، در حالی که سایر چربی ها، مانند فسفولیپیدها در سیگنال دهی سلولی و شناسایی سلول نقش دارند.
بخش سوم: انتقال مواد در عرض غشا
چه می آموزیم؟
تا اینجا آموختیم که غشا ساختاری بسیار مستحکم دارد و نفوذ هر ماده ای درون آن طبق برنامه ریزی و اصول مشخصی صورت می گیرد. حال بیایید ببینیم مواد از چه راه هایی وارد سلول می شوند؟
اسمز
قبل از ورود به انواع جابجایی در عرض غشا و برای درک بهتر آن باید ابتدا باید با مفهوم اسمز آشنا شویم. بیایید با مثال توضیح دهیم. ظرفی را تصور کنید که درون آن آب خالص وجود دارد. این ظرف به وسیله یک لایه نسبتاً نفوذپذیر به آب، به دو قسمت مساوی تقسیم می شود. درون یک قسمت، مقداری نمک ریخته و مخلوط می کنیم. اگر ظرف را به حال خود رها کنیم، به مرور آب از سمت بی نمک به سمت پر نمک جابجا می شود.
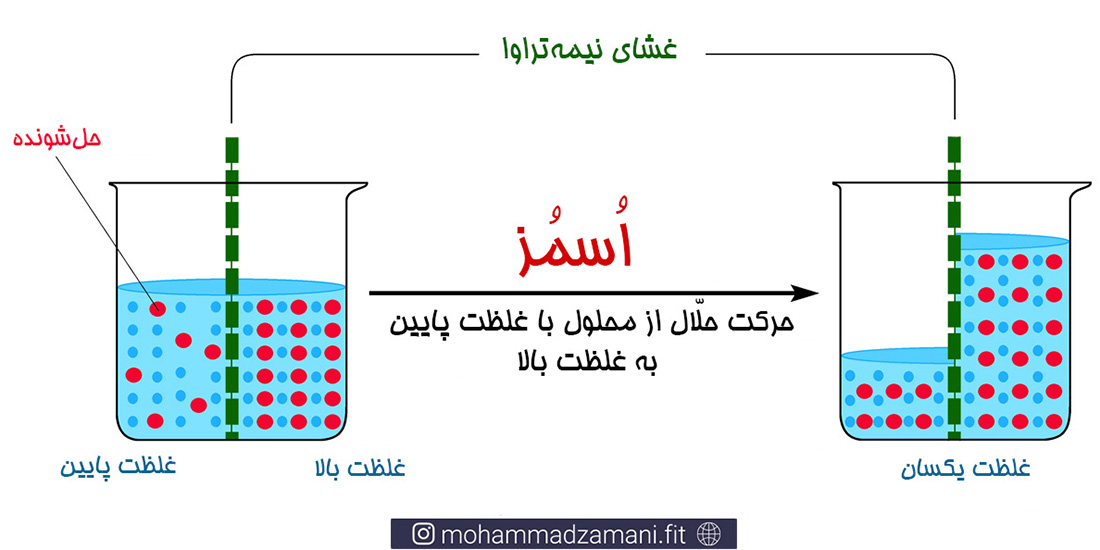
در واقع آب در جهتی جابجا می شود که غلظت مایع بیشتر است تا غلظت هر دو سمت را برابر سازد. نیرویی که باعث جابجایی آب در این شرایط می شود، با نام نیروی اسمزی شناخته می شود. این فرآیند درون بدن، اولین بار توسط ویلیام هوسون (William Hewson) در سال 1770 در سلول های قرمز خون کشف شد. آنان مشاهده کردند که زمانی که این سلول ها در محلول هاپوتونیک (محلول رقیق) قرار می گیرد، ورم می کند و زمانی که در محلول هایپرتونیک (محلول غلیظ) قرار می گیرد، جمع شده و چروکیده می شود (14).
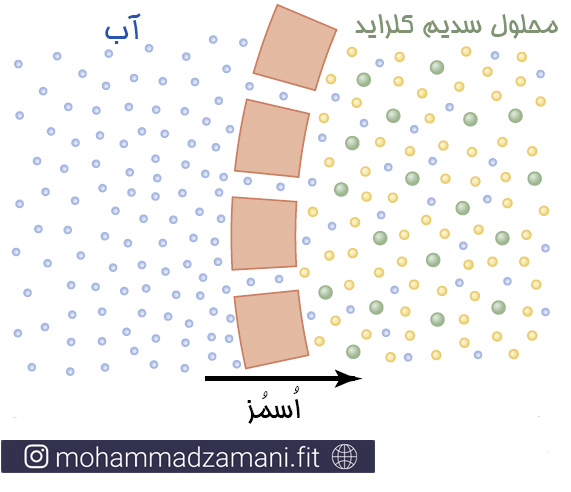
اسمز در غشای سلول زمانی که محلول سدیم کلراید در یک سمت غشا و آب در سمت دیگر غشا قرار می گیرد.
انواع جابجایی مواد در عرض غشا
از آنجایی که سلول، حتی به صورت ایزوله، یک موجود زنده است، با بستن همه درب ها به دنیای بیرون خود نمی تواند زنده بماند. بنابراین همواره راه هایی برای عبور مواد ضروری، چه به داخل و چه بیرون دارد. مواد به روش های مختلف در عرض غشا جابجا می شوند که در ادامه هر یک را توضیح می دهیم (15).
انتشار ساده
منظور از انتشار ساده، جابجایی مولکول ها یا یون ها در عرض غشا از طریق حفره های پروتئینی موجود در غشا یا از طریق فضای بین مولکولی، بدون تعامل با پروتئین های حامل در غشا است. میزان انتشار یک ماده به مقدار ماده در دسترس (شیب غلظتی)، سرعت حرکت ماده و تعداد و اندازه حفره های عبوری موجود بر روی غشا بستگی دارد.
انتشار ساده به دو شیوه اتفاق می افتد: 1) اگر ماده انتشار دهنده محلول در چربی باشد، از میان غشای دولایه لیپیدی عبور می کند؛ و 2) از طریق کانال های آبی که پروتئین های بزرگی هستند یک کانال عبور در عرض غشا ساخته اند.
میزان حلالیت پذیری در چربی، یکی از مهم ترین فاکتورهای تعیین کننده در انتشار ساده است. برای مثال حلالیت پذیری اکسیژن، نیتروژن، دی اکسید کربن و الکل ها در چربی، بسیار بالاست و همه این مواد می توانند مستقیما از غشا عبور کرده و به سمت دیگر ان منتشر شوند. میزان انتشار هر یک از این مواد نسبت مستقیمی با میزان حلالیت پذیری آن ها در چربی دارد.
اگرچه آب در غشای لیپیدی شدیداً حلال ناپذیر است، اما می تواند به آسانی از طریق کانالهای پروتئینی که به شکل سرتاسری عرض غشا را طی کرده اند، عبور کند. بیشتر سلول های بدن حاوی منافذ پروتئینی هستند که «آکواپورین» نامیده می شوند و به صورت انتخابی اجازه می دهند، آب به سرعت از عرض غشا عبور کند. آکواپورین ها تا حد زیادی ویژه هستند؛ به طوری که 13 نوع متفاوت از آکواپورین در سلول های مختلف پستانداران وجود دارد.
سرعت انتشار مولکول های آب از بیشتر غشاهای سلولی حیرت انگیز است. برای مثالف مقدار کل آبی که در غشای سلول های قرمز خون در هر دو جهت در ثانیه منتشر می شود، تقریباً 100 برابر کل حجم سلول است.
سایر مولکول های حلال ناپذیر در چربی، اگر در آب حلال پذیر بوده و به اندازه کافی کوچک باشند، می توانند از همین منافذ پروتئینی که آب عبور می کند، عبور کنند. ولی هر اندازه این ذرات بزرگتر شوند، به همان اندازه نفوذپذیری غشا به آن ها کاهش می یابد. جالب است بدانید که تعداد آکواپورین های غشایی در سلول ها ثابت نیست و بسته به شرایط فیزیولوژیک سلول می تواند تغییر کند.
کانال ها یا حفره های پروتئینی به وسیله دو ویژگی مهم دارند: اغلب نفوذپذیری انتخابی به مواد مشخص دارند؛ بیشتر کانال ها به وسیله دروازه هایی باز یا بسته می شوند. این دروازه ها با سیگنال های الکتریکی (کانال های ولتاژی (voltage-gated Channels)) یا مواد شیمیایی که به پروتئین کانال متصل می شود (کانال های لیگاندی (Ligand-gated Channels)) می توانند باز یا بسته شوند.
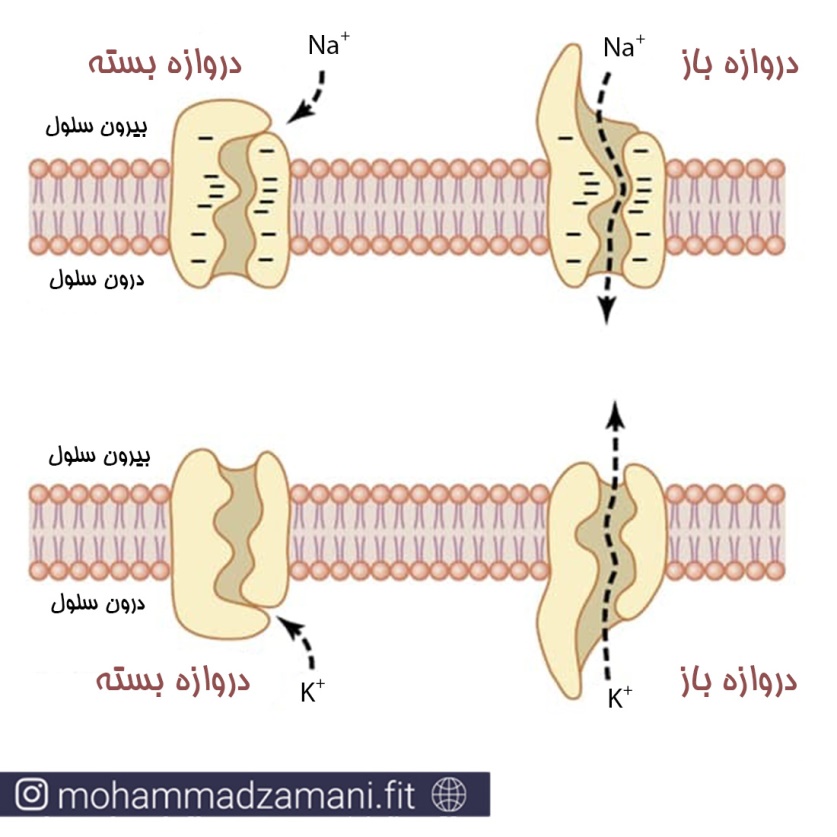
انتقال یون های سدیم و پتاسیم از طریق کانال های پروتئینی. در شکل می توانید تغییرات ساختاری پروتئین برای باز یا بسته شدن دریچه کانال را مشاهده فرمایید.
انتشار تسهیل شده
این روش جابجایی نیز مانند انتشار ساده است، با این تفاوت که در انتشار تسهیل شده، مواد به صورت هم انتقالی و با کمک یک حامل پروتئینی جابجا می شوند. انتشار ساده و تسهیل شده یک تفاوت اساسی با هم دارند. در انتشار ساده میزان انتقال نسبت مستقیم با غلظت ماده دارد؛ در حالی که در انتشار تسهیل شده، در هر شرایطی، یک میزان حداکثری برای انتشار وجود دارد که Vmax نامیده می شود. در شکل زیر می توانید تفاوت این دو را ببینید.
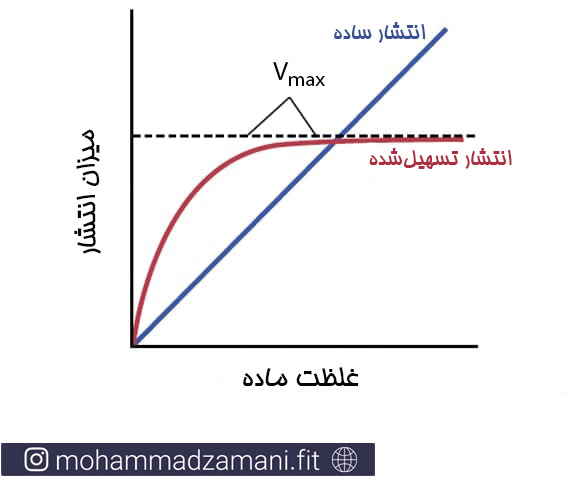
تأثیر غلظت ماده بر میزان انتشار از طریق غشا به وسیله انتشار ساده و انتشار تسهیل شده. این منحنی نشان می دهد که انتشار تسهیل شده دارای حد بیشینه ایست که Vmax نامیده می شود.
از جمله موادی که به روش انتشار تسهیل شده از غشا عبور می کنند، می توان گلوکز و بیشتر اسیدهای آمینه را نام برد. در مورد گلوکز، حداقل 14 نوع مختلف از انتقال دهنده آن (که GLUT نامیده می شوند) در سلول های مختلف وجود دارد. برخی از این GLUTها، مونوساکاریدهای دیگری را که ساختاری شبیه گلوکز دارند، مانندگالاکتوز و فروکتوز را نیز منتقل می کنند. یکی از این GLUTها (GLUT4) به وسیله انسولین فعال می شود که می تواند میزان انتشار تسهیل شده گلوکز را در بافت های حساس به انسولین، 10 تا 20 برابر افزایش دهد.
برای مطالعه بیشتر
غلظت املاح در دو سوی غشا
در ادامه بحث، با ارائه مثال و ذکر نمونه های مختلف، با انتقال مواد در عرض غشا بیشتر آشنا خواهیم شد. بیشترین موادی که در عرض غشا جابجا می شوند، املاح هستند. این موضوع برای حیات سلول بسیار اهمیت دارد. زیرا غلظت املاح در دوسوی غشا، تعیین کننده حجم سلول، بار الکتریکی آن و طبیعتاً بسیاری از عملکردهای طبیعی آن خواهد بود. بیایید نگاهی بیاندازیم به غلظت املاح و برخی از مواد در دو سوی غشا سلول استراحتی در انسان (16).
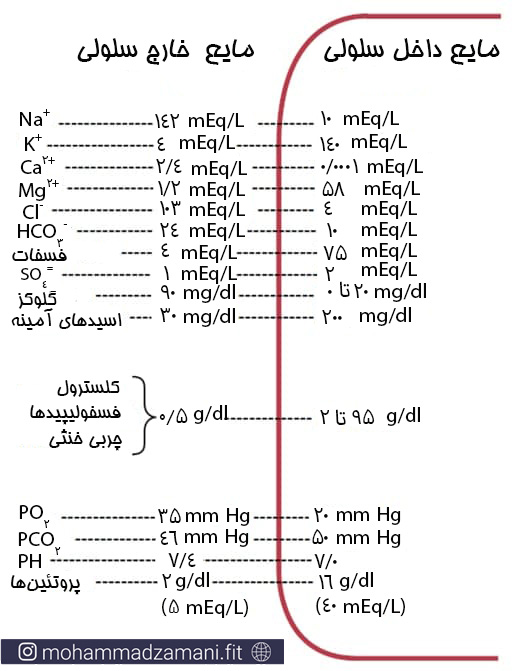
غلظت داخل سلولی و خارج از سلولی برخی از یون ها و مولکول ها
همانطور که مشاهده می کنید، غلظت برخی از مواد، مانند سدیم و کلر در بیرون از غشا و برخی مانند پتاسیم و منیزیوم در داخل غشا بیشتر است. در واقع شیب غلظتی برخی به سمت داخل و برخی به سمت خارج از غشا است. غشا در حالت عادی به این مواد نفوذ ناپذیر است تا اطمینان حاصل شود که غلظت این مواد در هر دو سمت به همین صورت باقی می ماند.
علاوه بر شیب غلظتی، نیروی مهم دیگری که باعث جابجایی غیرفعال مواد در عرض غشا می شود، نیروی الکتریکی است. درون سلول به دلیل داشتن مقادیر بالایی از مولکول های پروتئینی، دارای یون منفی قابل توجهی است و بیرون غشا در حالت استراحتی، مثبت تر از درون غشا است. بنابراین اگر غشا کاملا به این یون ها نفوذ پذیر شود، باز هم غلظت آن ها در دو سوی غشا برابر نخواهد شد و اندکی اختلاف وجود خواهد داشت.
عوامل اثرگذار بر میزان انتشار
بر اساس شواهد، مواد زیادی می توانند در عرض غشا منتشر شوند. اما میزان خالص این انتشار تحت تأثیر عوامل مختلف قرار می گیرد.
تفاوت غلظت در دو سوی غشا
شکل زیر، قسمت الف، یک غشا را نشان می دهد که غلظت مواد در خارج آن بیشتر از داخل آن است. میزان انتشار ماده به داخل غشا در این شرایط، نسبت مستقیم با غلظت مولکول ها در بیرون غشا دارد که تعیین می کند چه تعداد مولکول در خارج از غشا در هر ثانیه به هم برخور کنند.
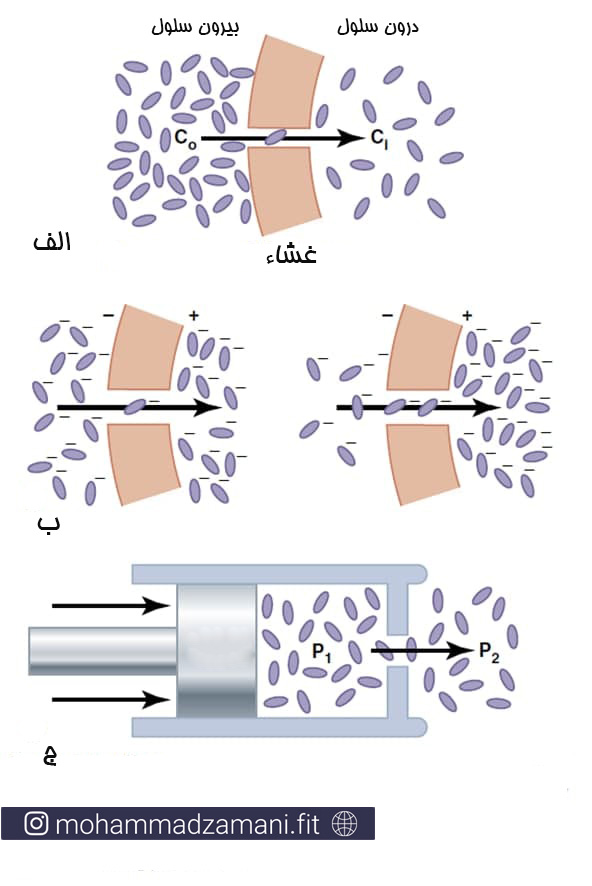
تأثیر اختلاف غلظت (الف)، اختلاف پتانسیل الکتریکی اثرگذار بر بار منفی (ب) و اختلاف فشار (ج) که باعث انتشار مولکول ها و یون ها از غشای سلول می شود.
پتانسیل الکتریکی غشا
اگر دو سوی غشا دارای اختلاف پتانسیل الکتریکی باشد، همانطور که در شکل بالا، قسمت ب مشخص شده است، بار الکتریکی یون ها سبب انتشار آن ها خواهد شد، حتی اگر نیروی نیروی کشش بار مخالف در خلاف جهت غلظت ماده باشد. برای مثل در شکل بالا، قسمت ب، علی رغم اینکه غلظت ماده در سمت داخل غشا بیشتر است، اما نیروی الکتریکی باعث جابجایی در جهت افزایش باز هم بیشتر غلظت یون در داخل غشا می شود.
اختلاف فشار در دوسوی غشا
گاهی اوقات، اختلاف فشار قابل توجهی بین دو سوی غشای اتفاق می افتد. این اختلاف فشار، برای مثال در غشای مویرگ های خونی بافت های بدن ایجاد می شود. به طوری که فشار درون مویرگ حدود 20 میلی متر جیوه بیشتر از خارج آن است.
فشار در واقع برآیند ضربات مولکول های مختلف بر واحد مشخصی از سطح در یک لحظه معین است. در بیشتر موارد، این شرایط ناشی از تعداد بالاتر مولکول ها در یک سمت است که باعث افزایش تعداد ضربات آن ها به هم در اثر حرکات تصادفی می شود. این اثر در شکل بالا، بخش ج نشان داده شده است.
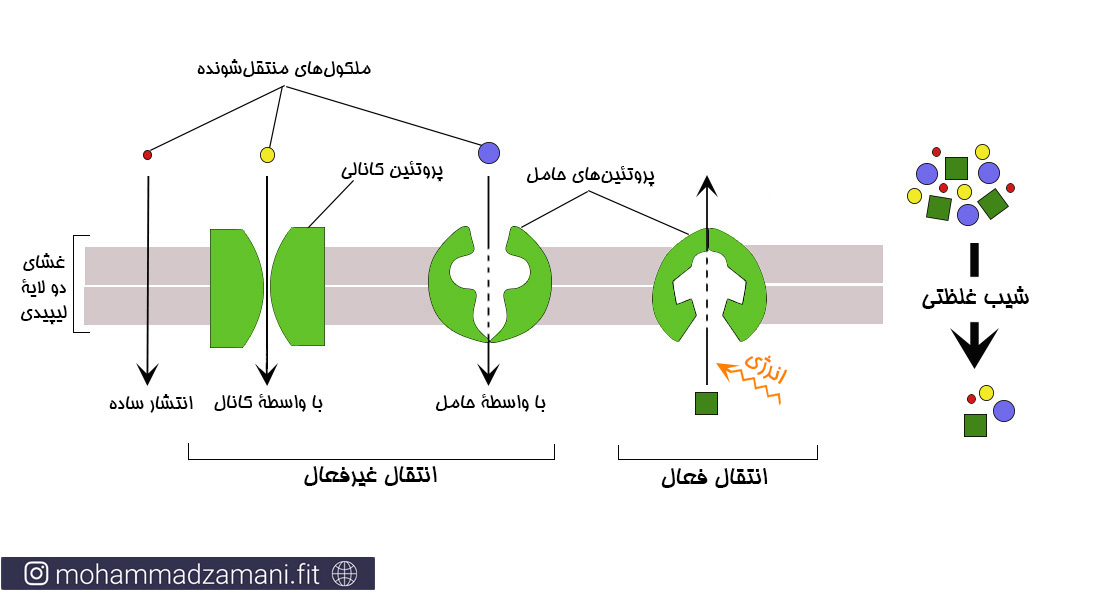
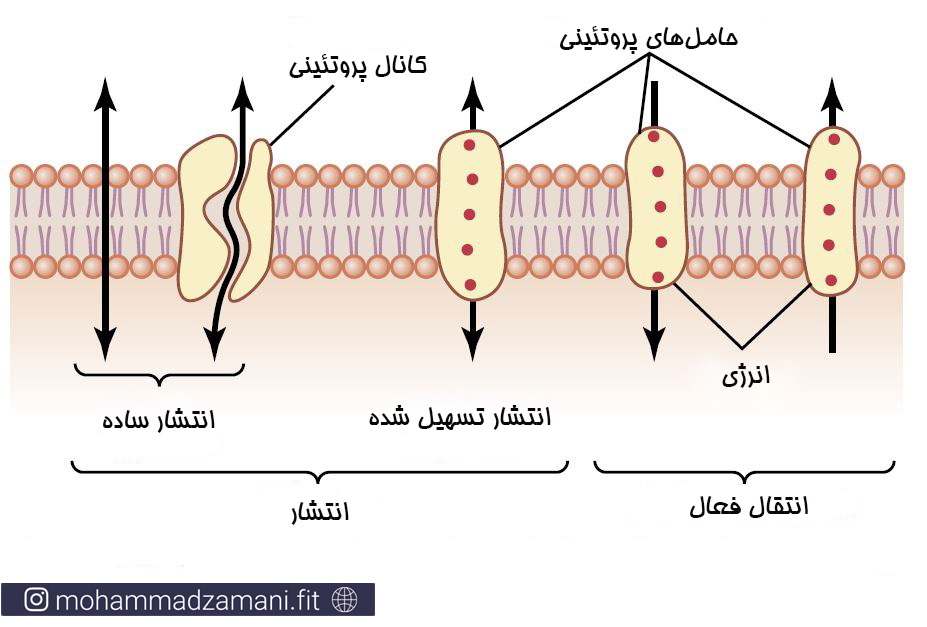
انواع جابجایی مواد از طریق غشا، با دو دسته بندی متفاوت؛ بالا (15) و پایین(16)
انتقال فعال
گاهی اوقات غلظت بالایی از یک ماده در مایع درون سلولی نیاز است، آن هم در شرایطی که غلظت همان ماده در مایع خارج سلولی بسیار اندک است. این شرایط خیلی هم فرضی نیست و مثالی از آن، یون پتاسیم است. البته برعکس این شرایط نیز وجود دارد که غلظت کم ماده در داخل سلول نیاز باشد و هم زمان غلظت آن در خارج سلول بسیار بالا باشد؛ مانند یون سدیم. در چنین شرایطی، باید نیرویی وجود داشته باشد که برخلاف شیب غلظتی، باعث حرکت یون های پتاسیم به داخل و حرکت یون های سدیم به خارج از سلول شود. زمانی که غشا سلول، مواد را در خلاف جهت شیب غلظتی، شیب الکتریکی یا شیب فشاری آن ها انتقال می دهد، به این نوع جابجایی، «انتقال فعال» می گویند. مثال هایی از موادی که با انتقال فعال در عرض غشا جابجا می شوند، یون های سدیم، پتاسیم، کلسیم، آهن، هیدروژن، کلراید، یداید و اورات، انواعی از قندها و بیشتر آمینو اسیدها است.
انتقال فعال اولیه و ثانویه
انتقال فعال بر اساس منبع انرژی مورد استفاده به دو نوع انتقال فعال اولیه و انتقال فعال ثانویه تقسیم می شود. در انتقال فعال اولیه، انرژی مورد نیاز، مستقیما از تجزیه ATP یا دیگر ترکیبات دارای فسفات پرانرژی حاصل می شود. در انتقال فعال ثانویه، انرژی به طور ثانویه از اختلاف غلظت یونی مواد بین دو طرف غشای سلولی که ذخیره شده است، که در اصل توسط حمل و نقل فعال اولیه ایجاد شده بود، حاصل می شود. در هر دو مورد، انتقال به «پروتئین های حامل» وابسته است که بطور سرتاسری عرض غشا را پیموده اند. در انتقال فعال، عملکرد پروتئین های حامل با انتشار تسهیل شده متفاوت است. حامل های انتقال فعال قابلیت این را دارند که انرژی را به ماده جابجا شوند منتقل کنند تا بتواند در خلاف جهت شیب الکتروشیمیایی خود جابجا شود.
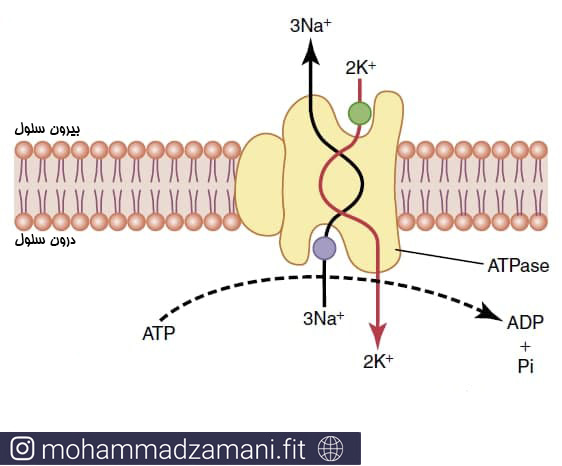
پمپ سدیم-پتاسیمی، نمونه ای از انتقال فعال اولیه (16)
مثالی از انتقال دهنده اولیه، پمپ سدیم-پتاسیم است که سدیم را به بیرون و پتاسیم را به داخل سلول انتقال می دهد. این پمپ مسئول حفظ غلظت یون ها در دو سوی غشا و نیز حفظ پتانسیل الکتریکی منفی تر در سمت داخلی غشا است. در برخی از سلول ها، مانند سلول های عصبی که از نظر الکتریکی فعال هستند، 60 تا 70 درصد انرژی سلول صرف پمپ کردن سدیم به بیرون و پتاسیم به داخل سلول می شود.
انتقال فعال ثانویه به صورت هم انتقالی و انتقال متقابل انجام می شود. زمانی که یون های سدیم به وسیله انتقال فعال اولیه به بیرون از سلول منتقل می شود، شیب غلظتی یون سدیم بین دوسوی غشا تشدید می شود، بصورتی که غلظت سدیم در بیرون غشا، بازهم بیشتر و در داخل غشا، بازهم کمتر می شود. این شیب غلظتی، نمایانگر یک مخزن انرژی است، زیرا سدیم مازاد در خارج از سلول، همواره تلاش می کند تا به داخل منتشر شود. تحت شرایط مناسب، این انرژی انتشار سدیم، می تواند مواد دیگر را نیز همراه خود به درون سلول بکشاند. این پدیده هم انتقالی نامیده می شود که یکی از انواع انتقال فعال ثانویه است.
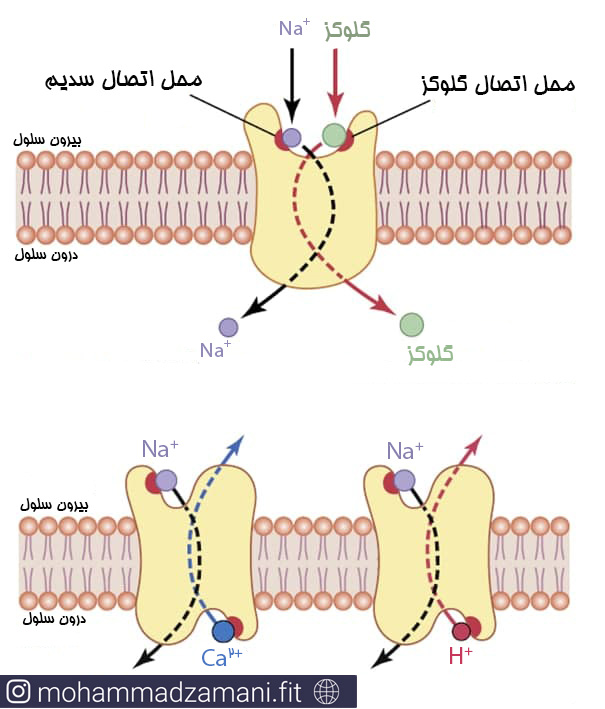
هم انتقالی سدیم و گلوکز (بالا) و انتقال متقابل سدیم و هیدروژن (پایین) در عرض غشا (16)
در انتقال متقابل که نوع دوم از انتقال فعال ثانویه است، یون سدیم مجدداً تلاش می کند تا در جهت شیب غلظتی خود منتشر شود. اما این بار ماده ای که قرار است منتقل شود در داخل سلول قرار دارد و به بیرون منتقل می شود. بنابراین، یون سدیم به پروتئین حامل، در قسمت بیرونی آن متصل شده و ماده ای که می خواهد جابجا شود به قسمت داخلی آن می چسبد. زمانی که هر دو ماده در دوسمت مخالف به پروتئین حامل بچسبد، حامل تغییر موقعیت داده و همزمان با انتقال سدیم به داخل، ماده دیگر را به خارج منتقل می کند (16).
این مقاله چه کمکی به ما کرد؟
در این مقاله ما با غشای سلول به عنوان یک میکروارگانل فعال در سلول آشنا شدیم. ساختار آن را به صورت اجمالی بررسی کردیم و نقش هریک از بخش های سازنده آن را دیدیم. همچنین مهم ترین کارکرد غشا، که نفوذ انتخابی است بررسی کردیم و دیدیم که مواد برای عبود از غشا چگونه عمل می کنند.
منابع
- https://scholar.google.com/scholar?hl=en&as_sdt=0,5&q=Origin+and+development+of+the+microscope+the+royal+microscopical+society
- https://ia800708.us.archive.org/view_archive.php?archive=/28/items/crossref-pre-1923-scholarly-works/10.1111%252Fj.1365-2133.1922.tb11337.x.zip&file=10.1111%252Fj.1365-2818.1915.tb04962.x.pdf
- https://www.researchgate.net/profile/Avraham-Hirschberg/publication/257439298_Schlieren_and_Shadowgraph_Techniques_Visualizing_Phenomena_in_Transparent_Media_GS_Settles_Springer-Verlag_Berlin_Germany_2001/links/59eef068aca272029ddf9619/Schlieren-and-Shadowgraph-Techniques-Visualizing-Phenomena-in-Transparent-Media-GS-Settles-Springer-Verlag-Berlin-Germany-2001.pdf
- https://books.google.com/books?hl=en&lr=&id=0DYXk_9XX38C&oi=fnd&pg=PA5&dq=Micrographia+or+some+physiological+descriptions+of+minute+bodies+made+by+magnifying+glasses.&ots=YYnk5dW__R&sig=gq0x5_NOB_VOrRQBXcHE15nSJGE#v=onepage&q=Micrographia%20or%20some%20physiological%20descriptions%20of%20minute%20bodies%20made%20by%20magnifying%20glasses.&f=false
- https://journals.biologists.com/jcs/article/s3-93/22/157/64031/The-Cell-Theory-A-Restatement-History-and-Critique
- https://books.google.com/books?hl=en&lr=&id=UinSjEa6PgwC&oi=fnd&pg=PA388&dq=Philosophie+Zoologique.+Paris%3B+1809&ots=opgi4XM3_q&sig=gizi-xoolzGxBs01K69OG_Kkv7M#v=onepage&q=Philosophie%20Zoologique.%20Paris%3B%201809&f=false
- https://books.google.com/books?hl=en&lr=&id=t_biw80LgjwC&oi=fnd&pg=PP1&dq=Plant+cell+biology+from+astronomy+to+zoology&ots=pWQVzYM9Vl&sig=I7bP9YrQs_fBz94M3mtZh3M8WLU#v=onepage&q=Plant%20cell%20biology%20from%20astronomy%20to%20zoology&f=false
- https://academic.oup.com/aob/article/106/4/vi/108266?login=true
- https://books.google.com/books?hl=en&lr=&id=uOxjtTE9koUC&oi=fnd&pg=PP1&dq=Membrane+permeability:+100+years+since+Ernest+Overton.&ots=MRifopV70q&sig=XDQNl1_1UTqNkGGxntZB6OqlHnE#v=onepage&q=Membrane%20permeability%3A%20100%20years%20since%20Ernest%20Overton.&f=false
- https://journals.sagepub.com/doi/pdf/10.1177/003591576706011P175
- https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2130960/
- https://www.science.org/doi/abs/10.1126/science.175.4023.720
- https://www.thoughtco.com/glycoprotein-definition-and-function-4134331
- https://www.sciencedirect.com/science/article/abs/pii/000930849390065B
- https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7182109/
- https://www.amazon.com/Guyton-Hall-Textbook-Medical-Physiology/dp/0323597122

